| 加齢研フェロー | 安井 明 |

加齢研フェロー
平成28年4月より加齢研で新たに「フェロー」職が創設されました。この「加齢研フェロー」職は非常勤で、科研費や寄付金などの外部資金を運用しますが、科研費等の申請の際の職名として登録され、加齢研共同研究の受入教員を任され、教授会へのオブザーバー参加を許されています。事務上は特定の研究分野(分子腫瘍学)に属します。この職は加齢研の研究者や研究活動の可能性を広げることが期待されます。
研究グループ

研究活動
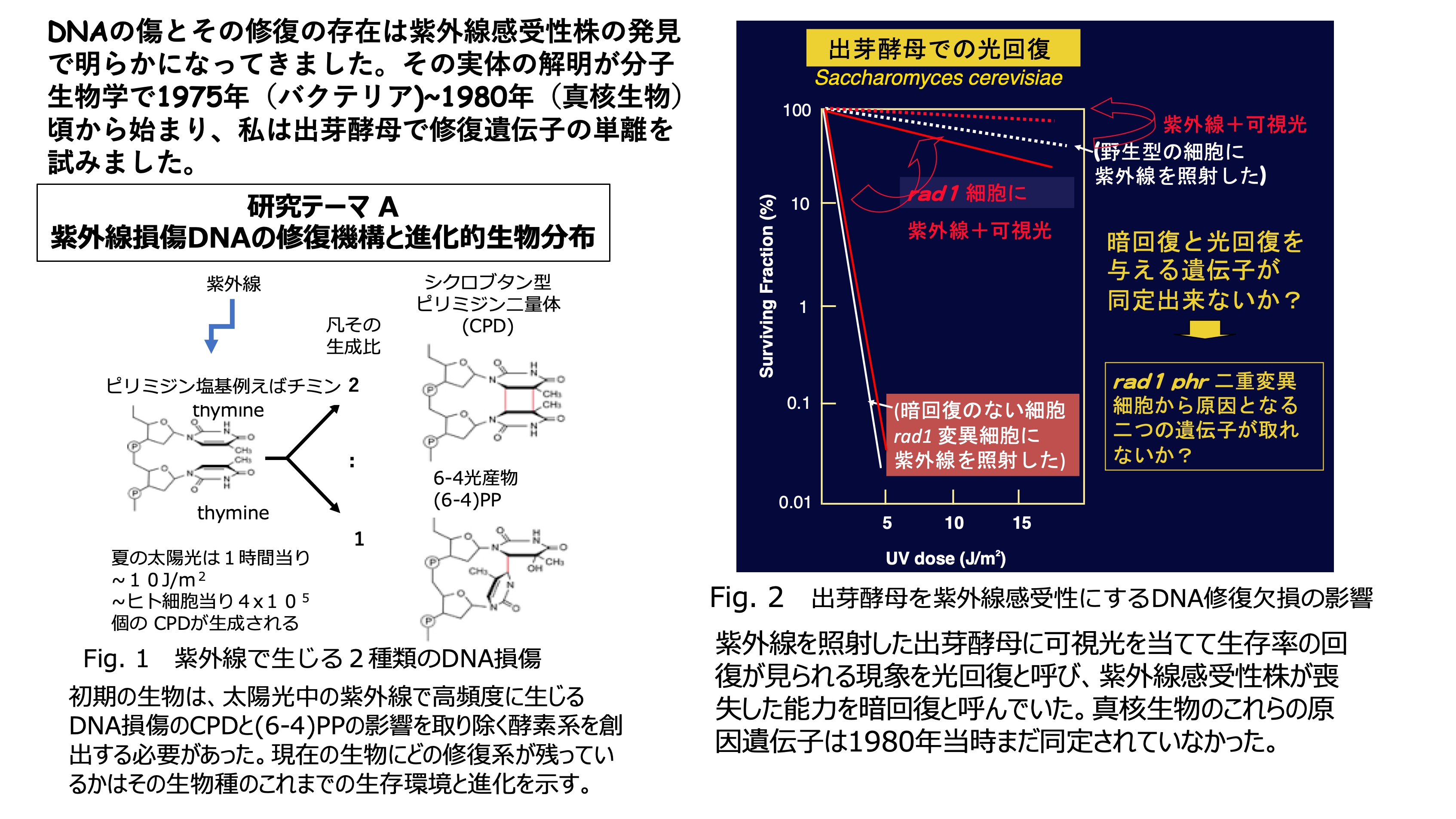
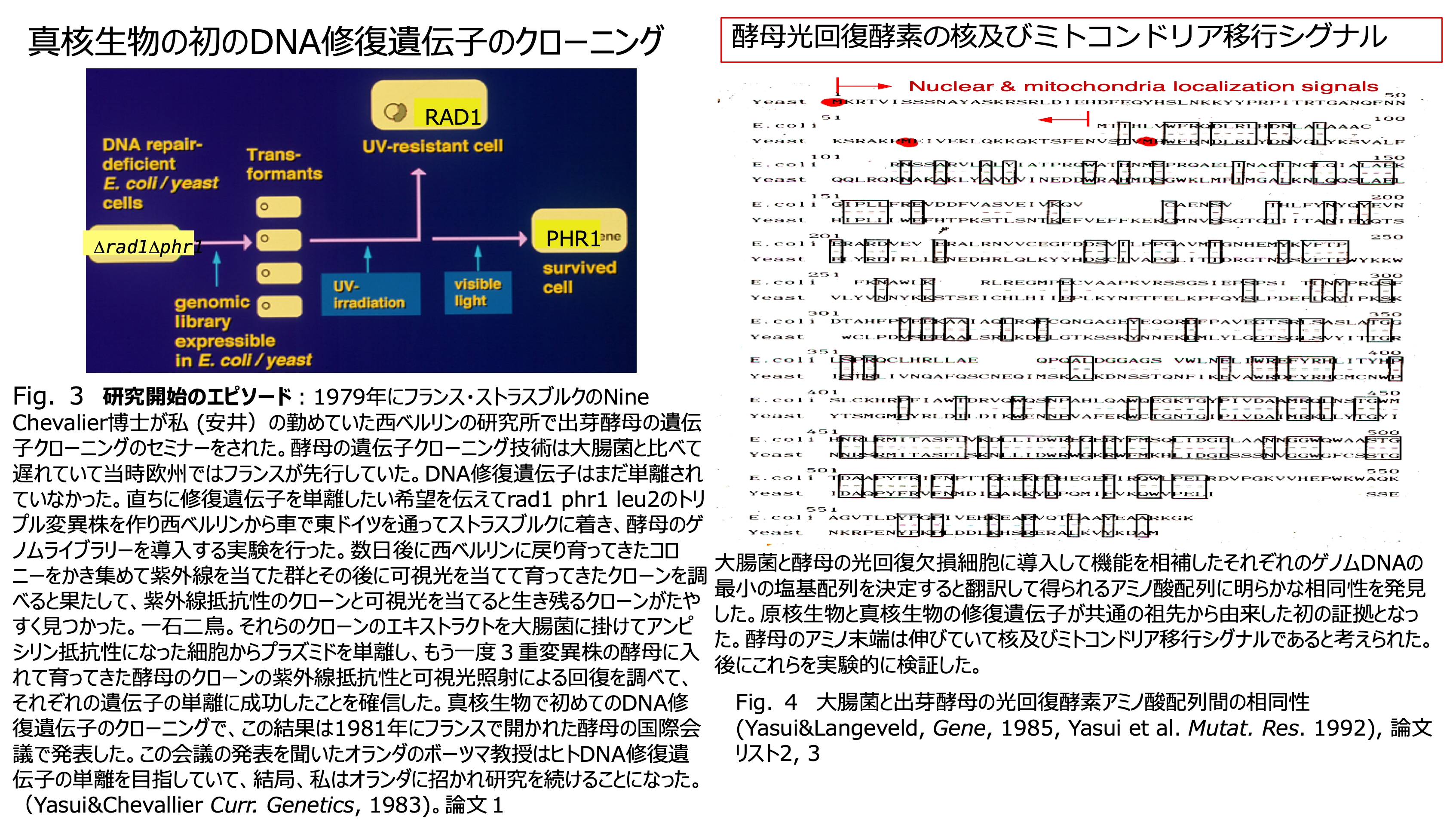
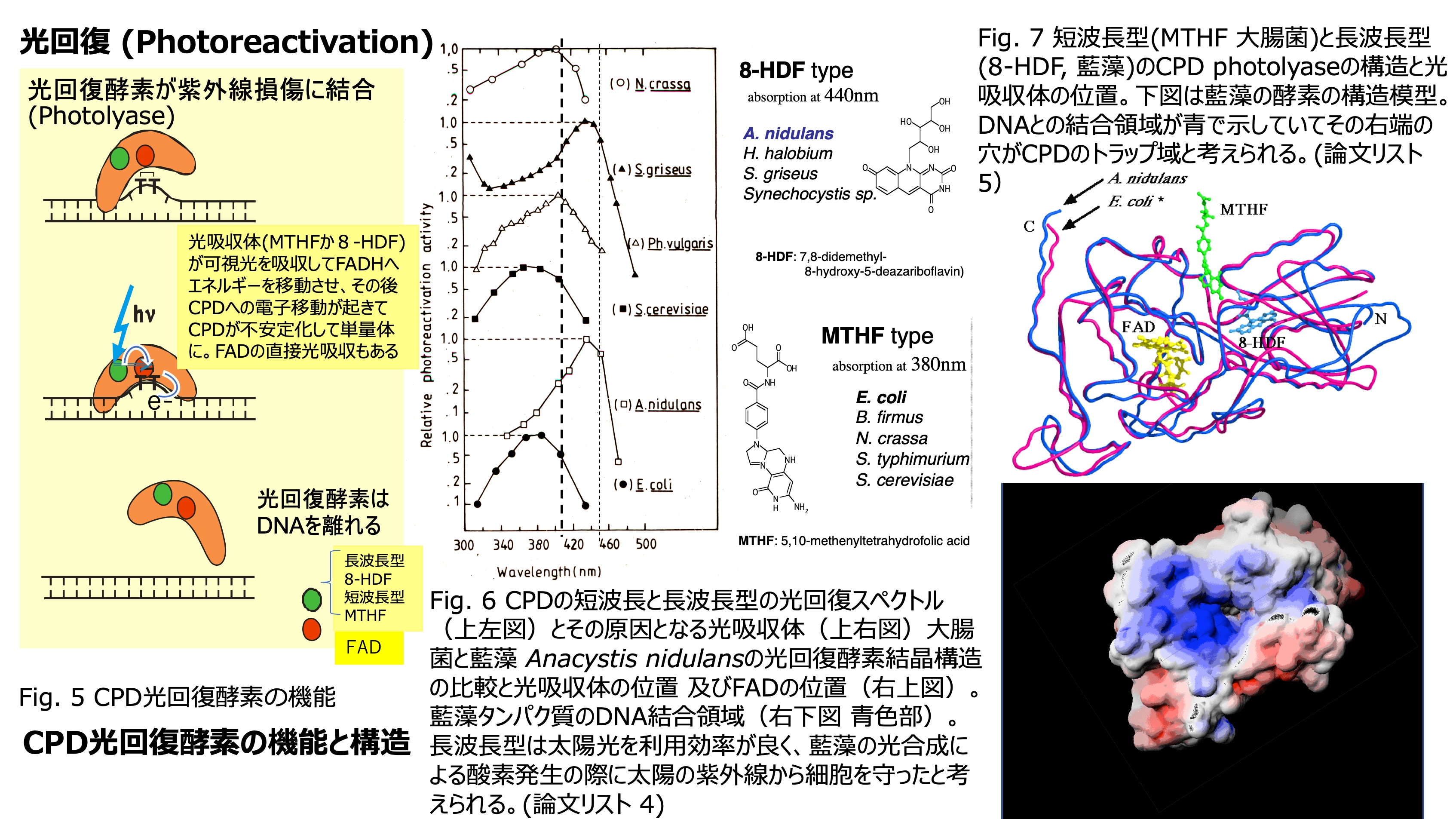
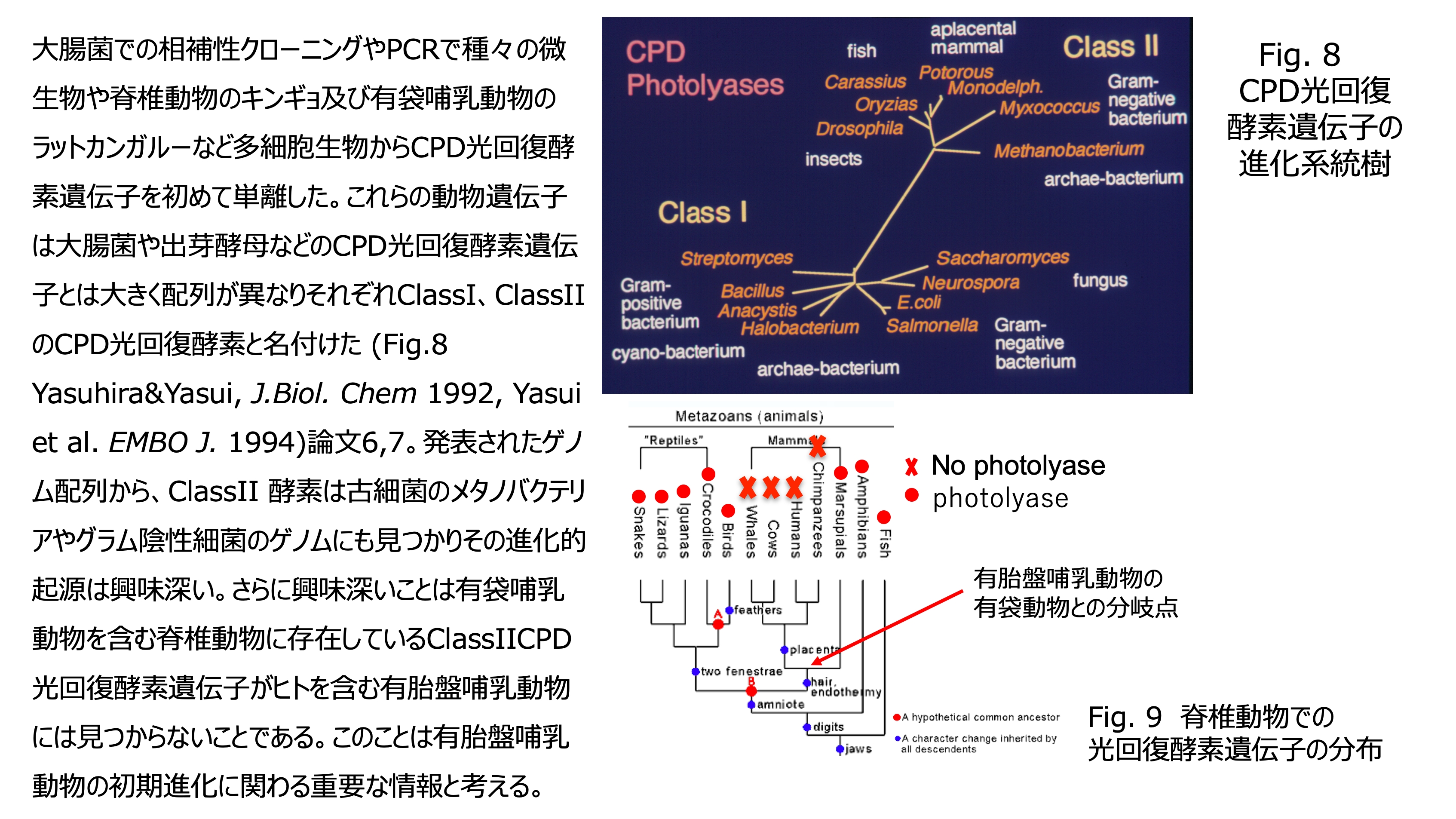
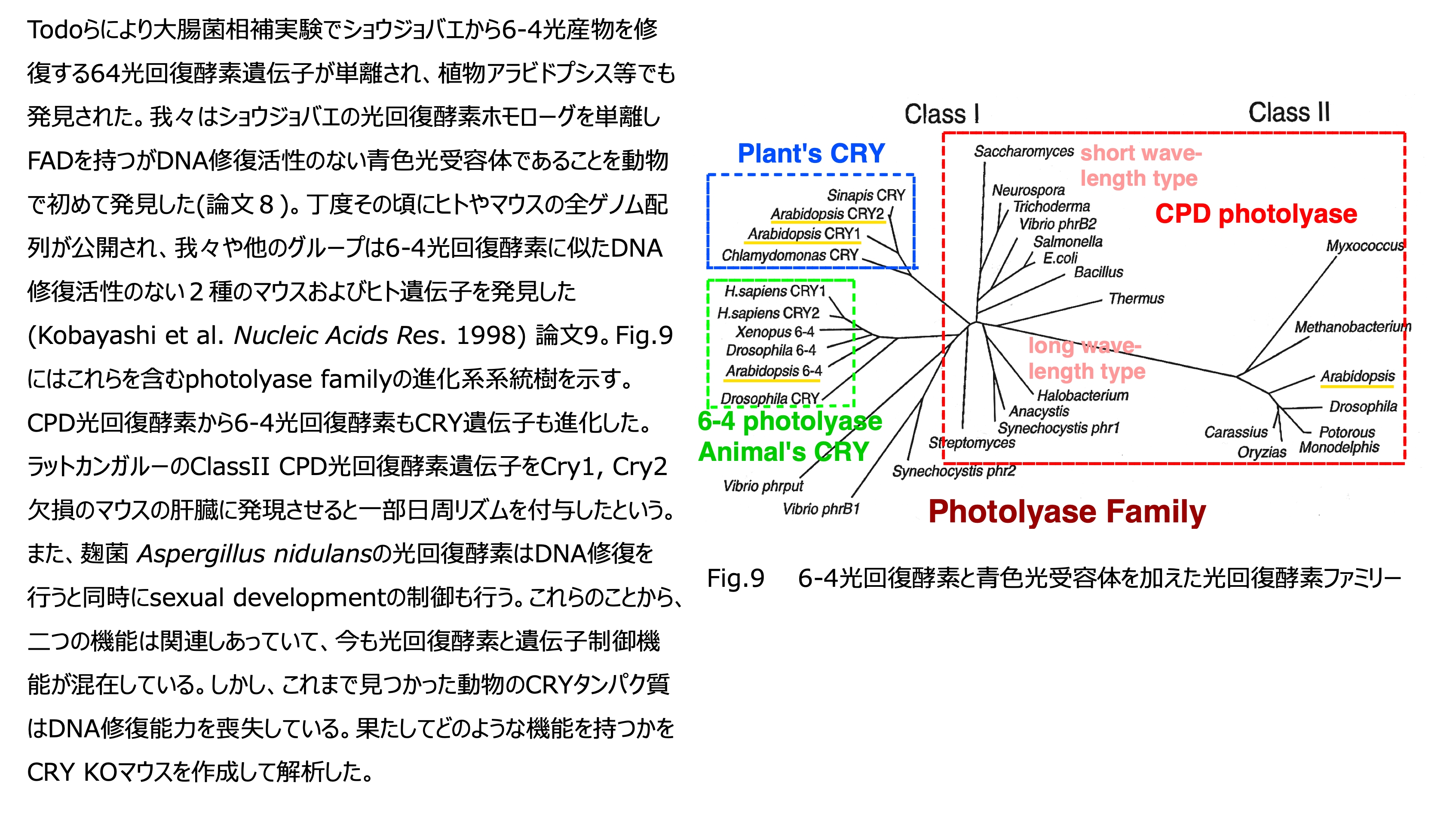

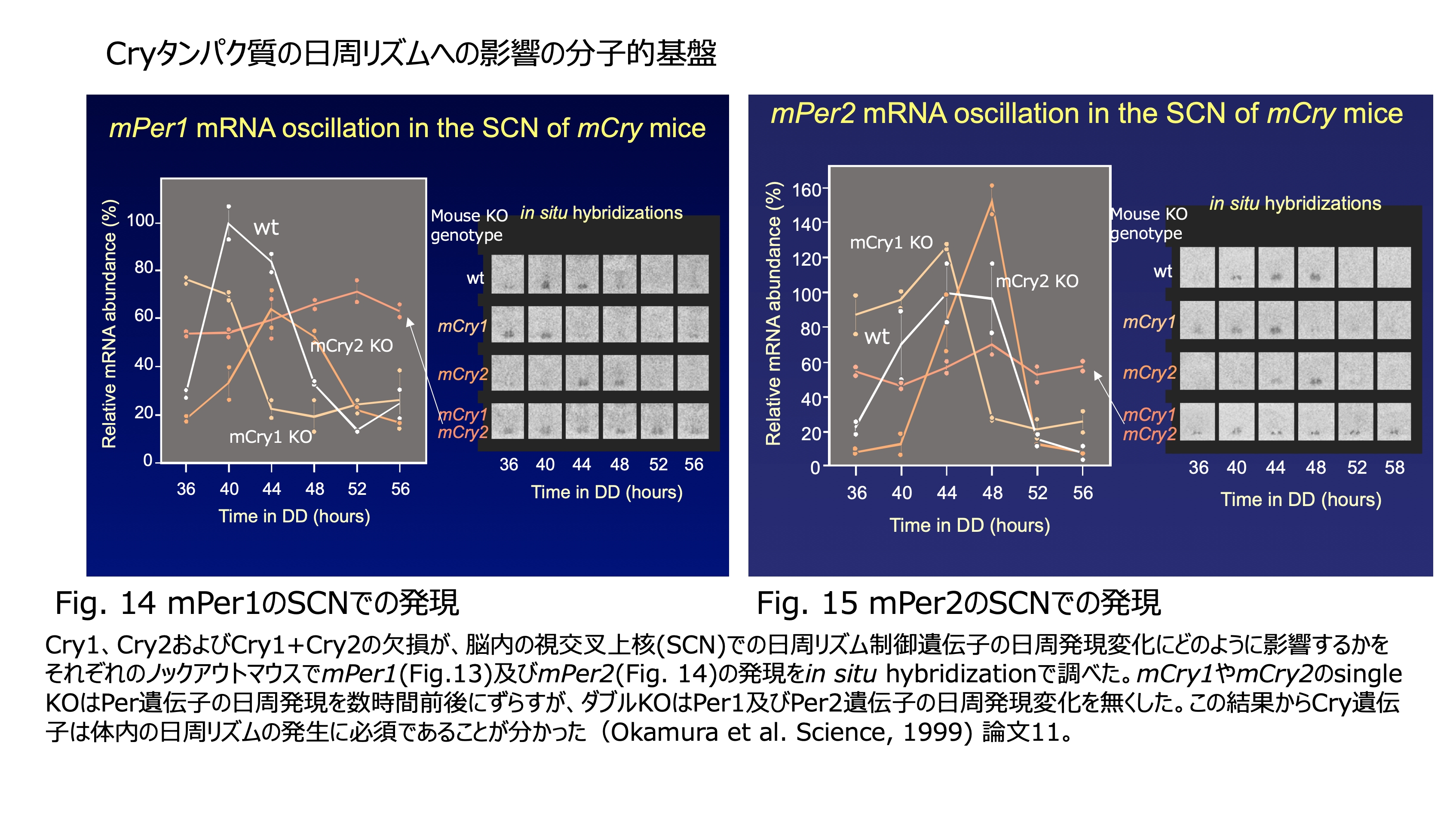
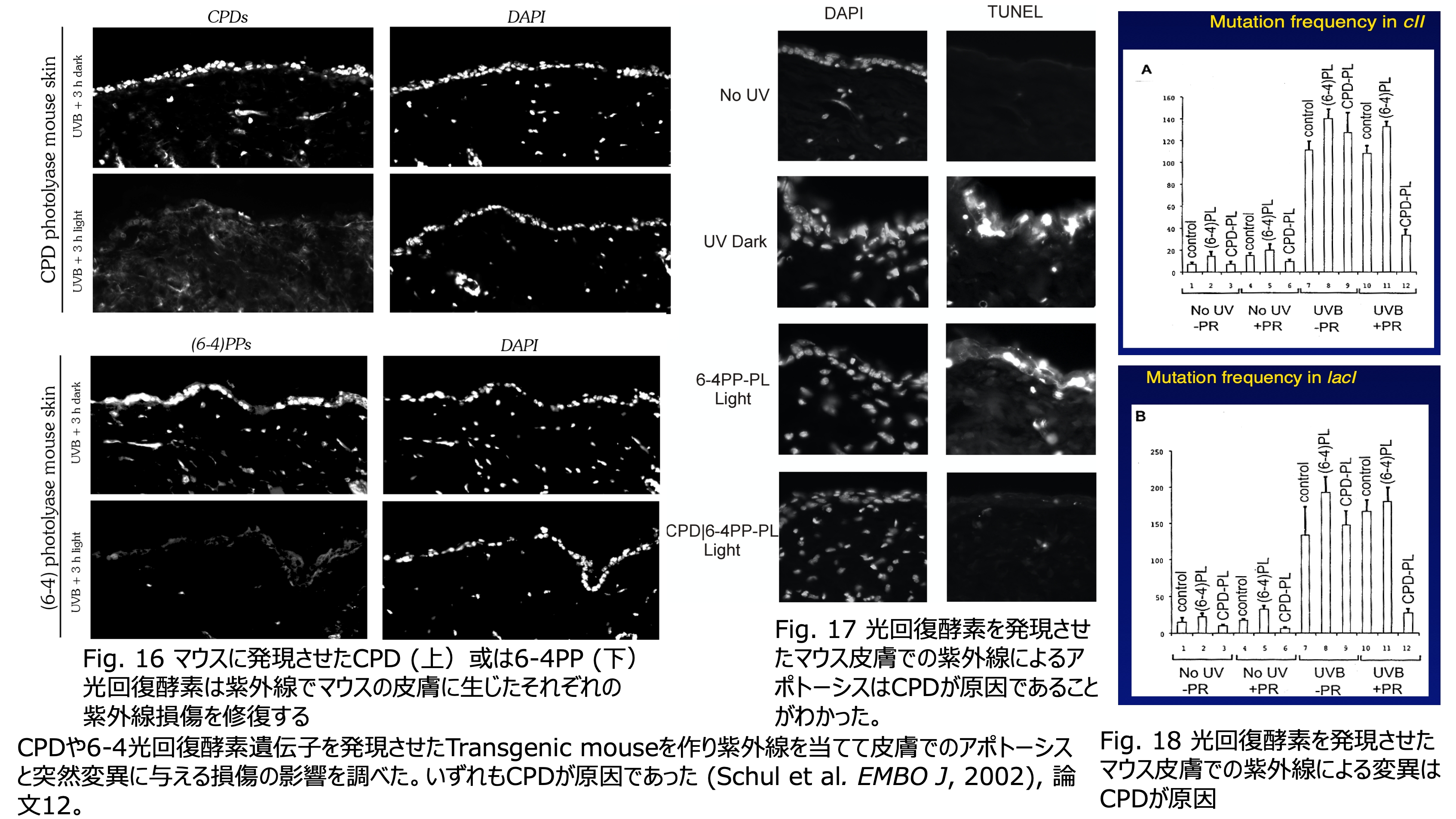
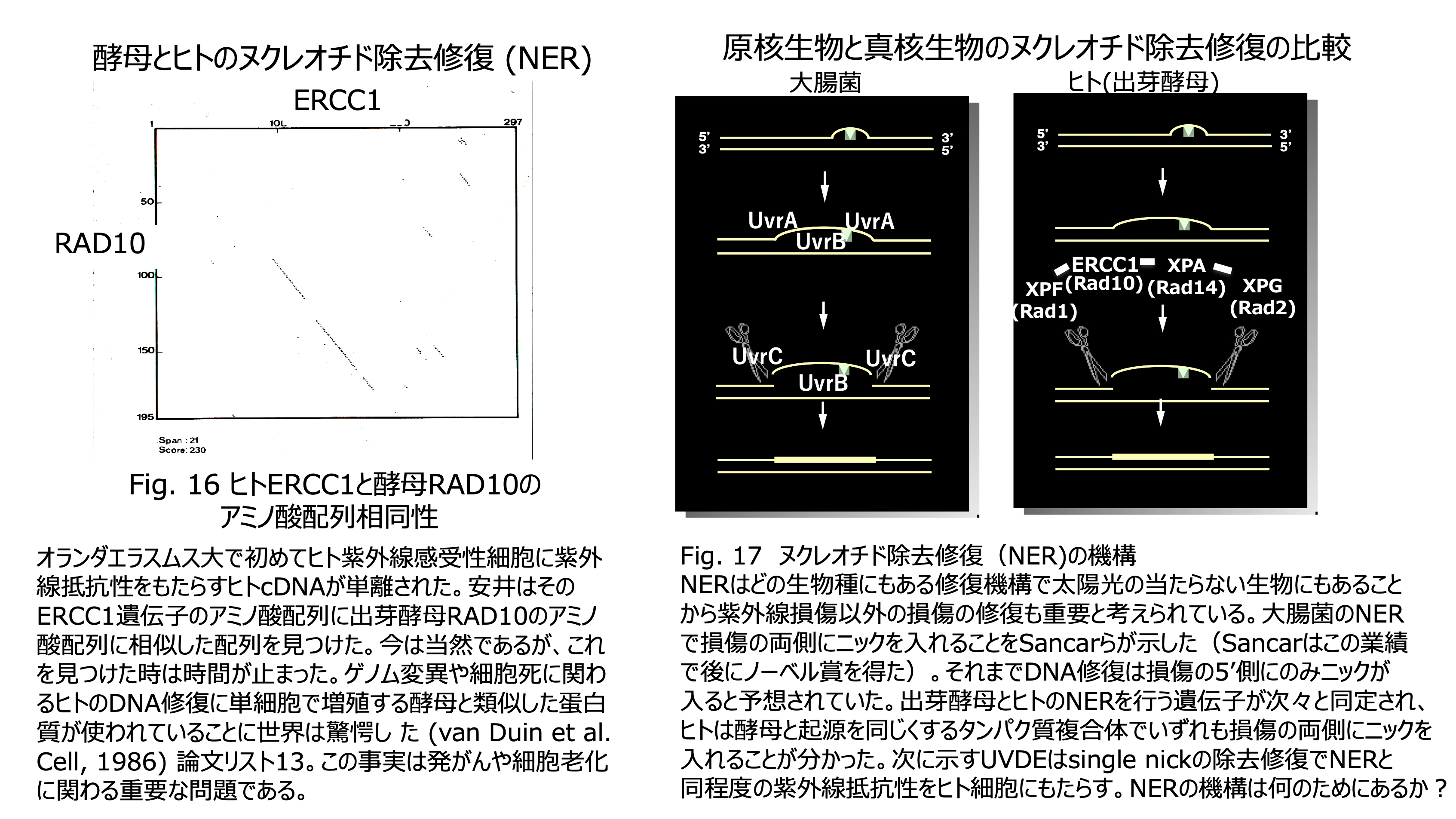
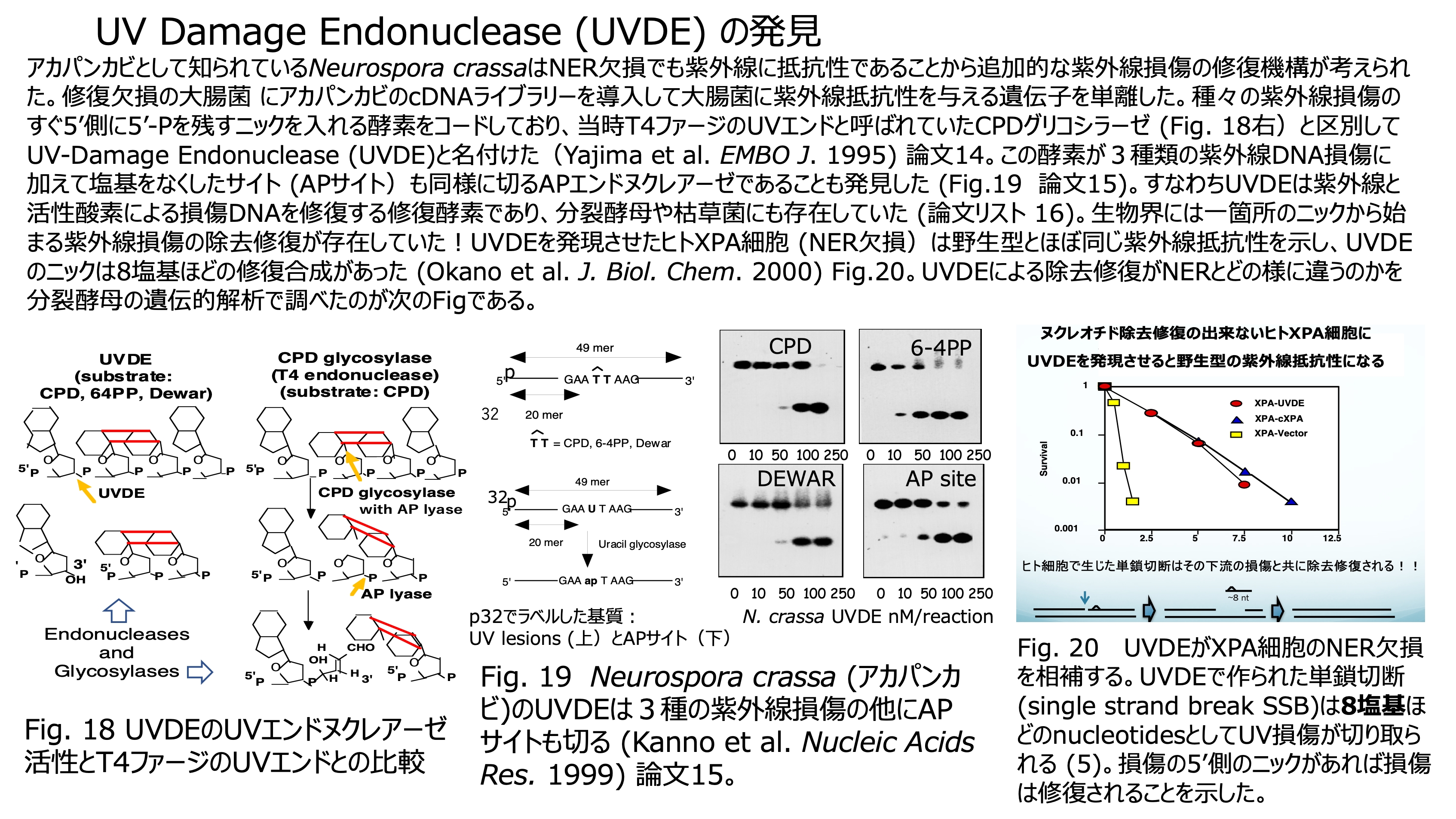
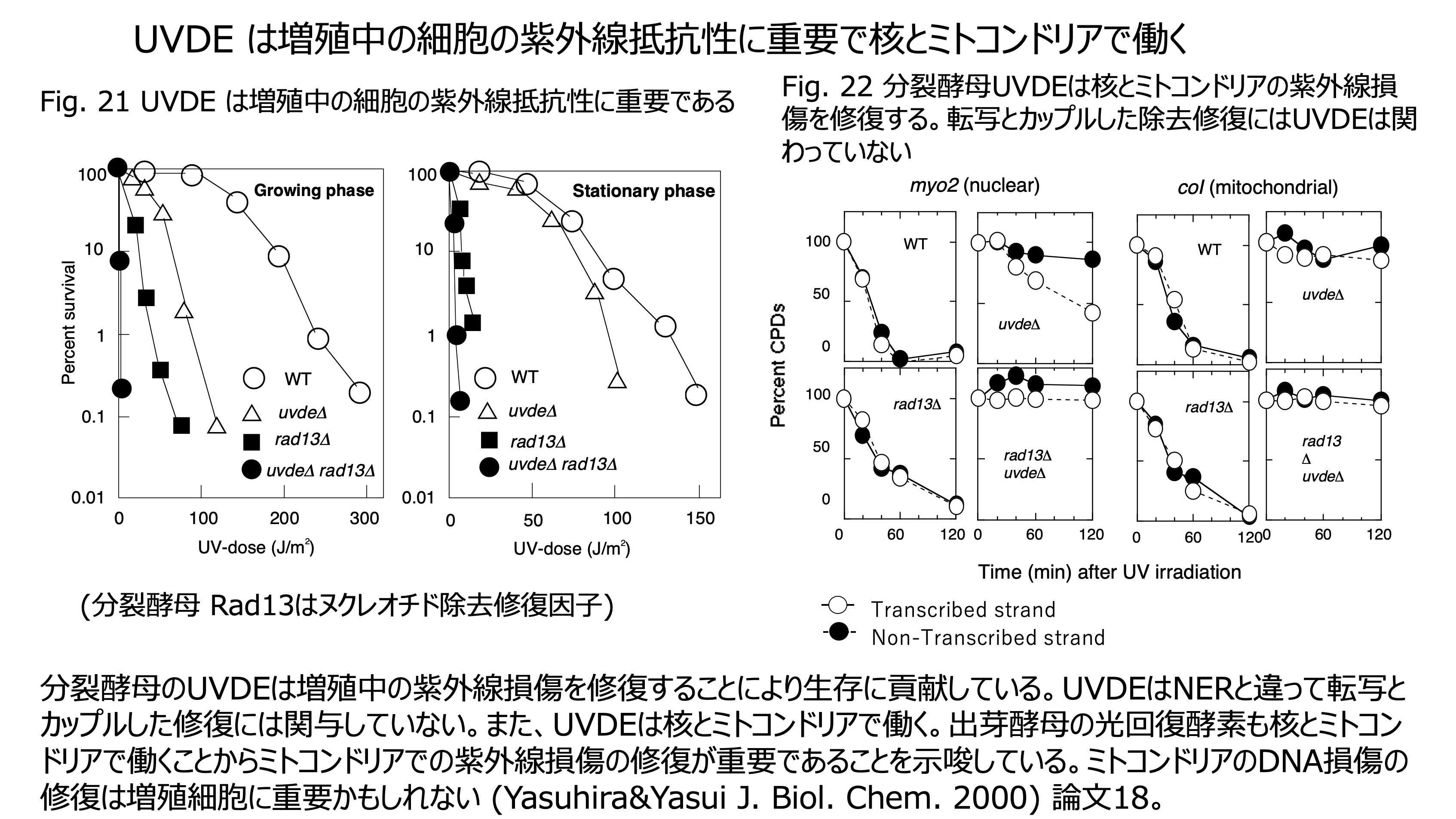
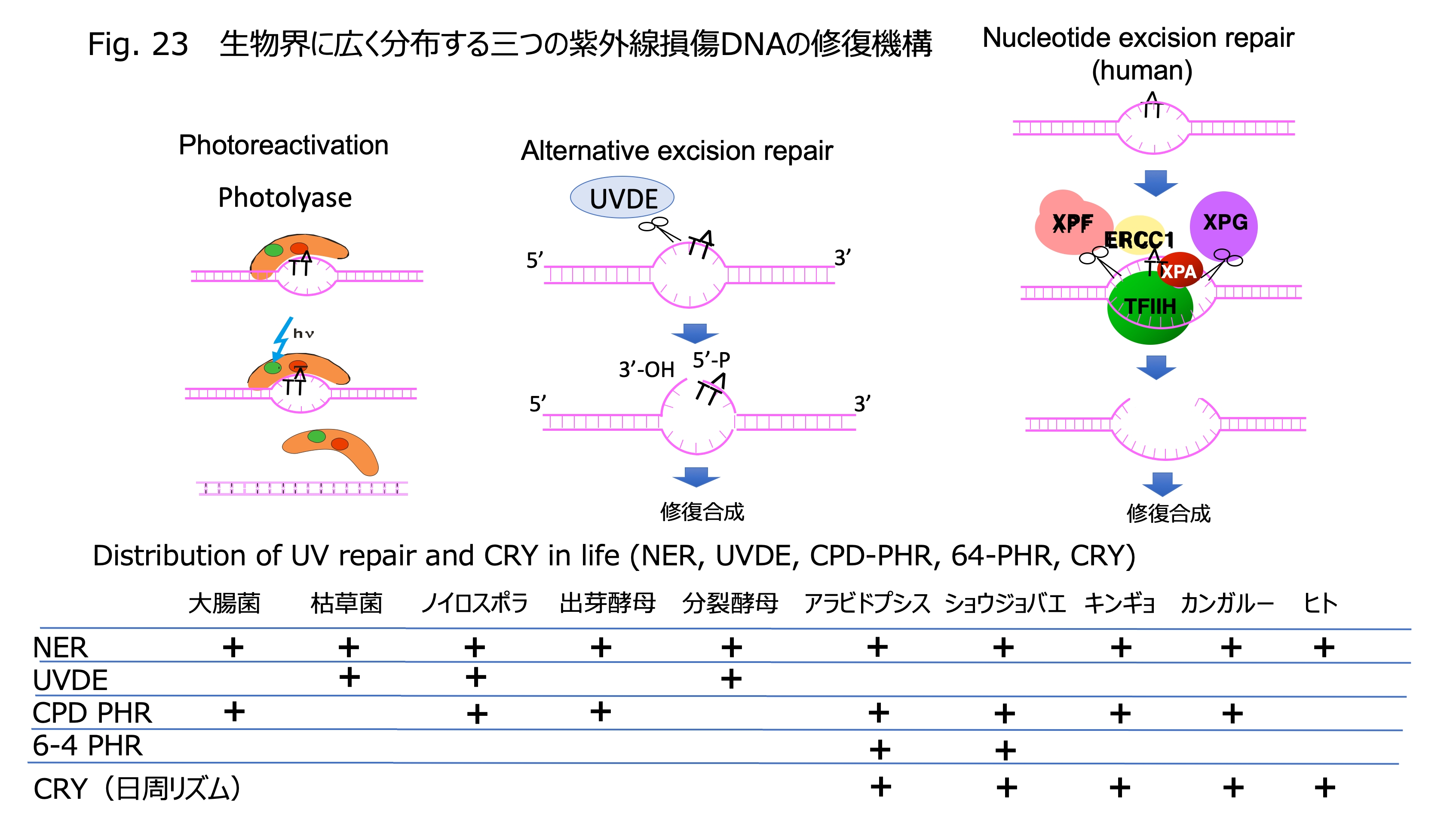
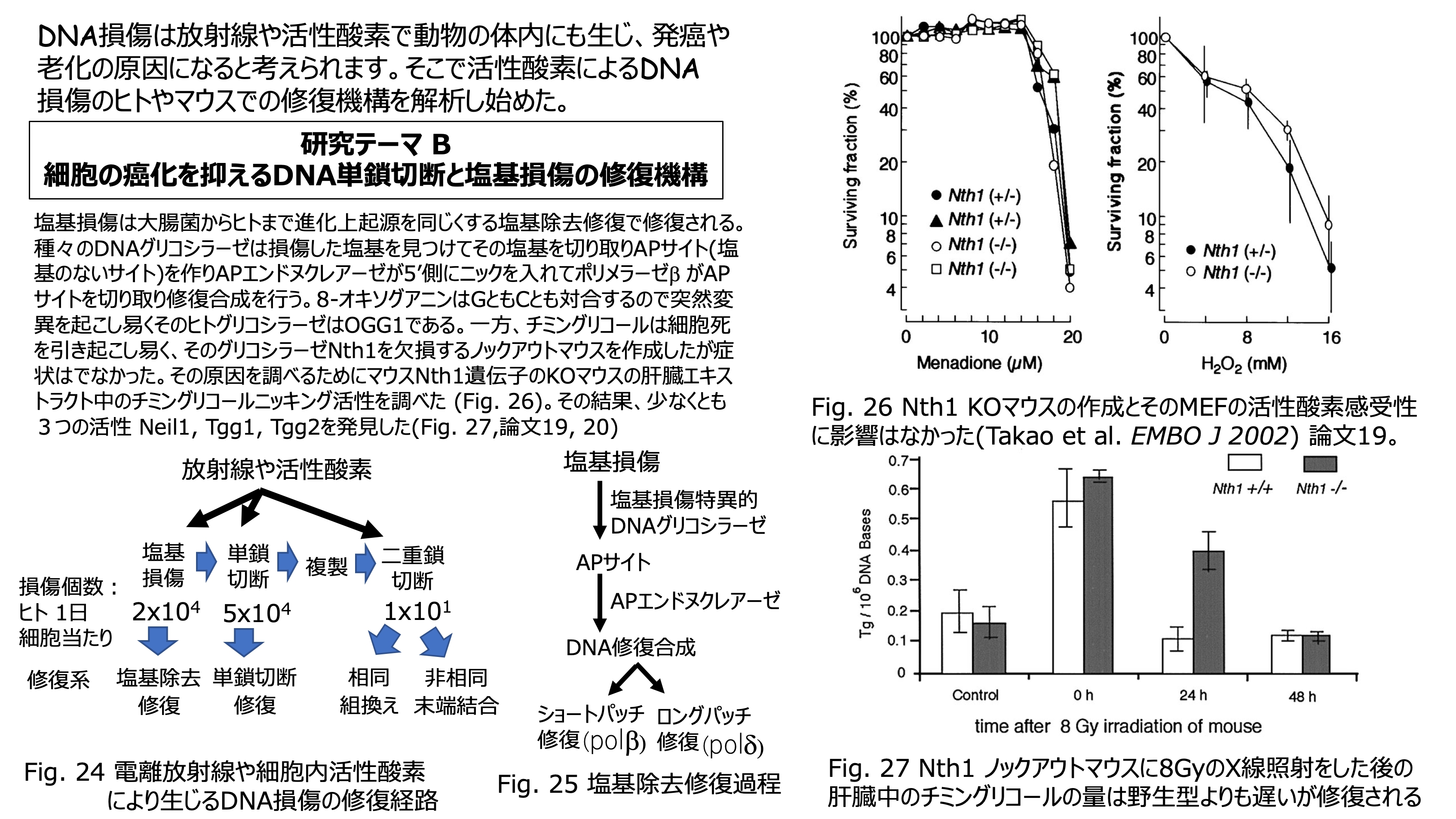
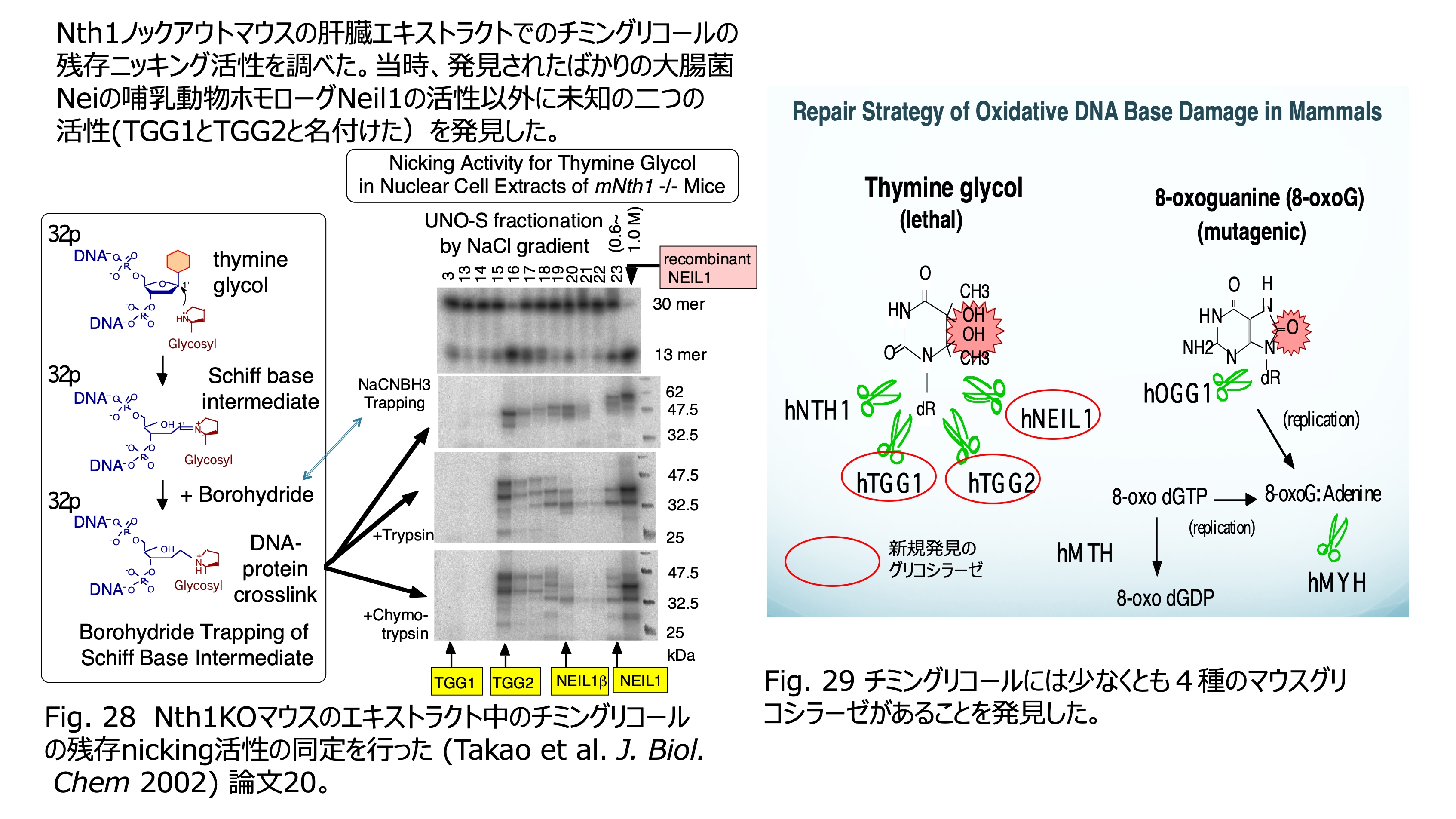
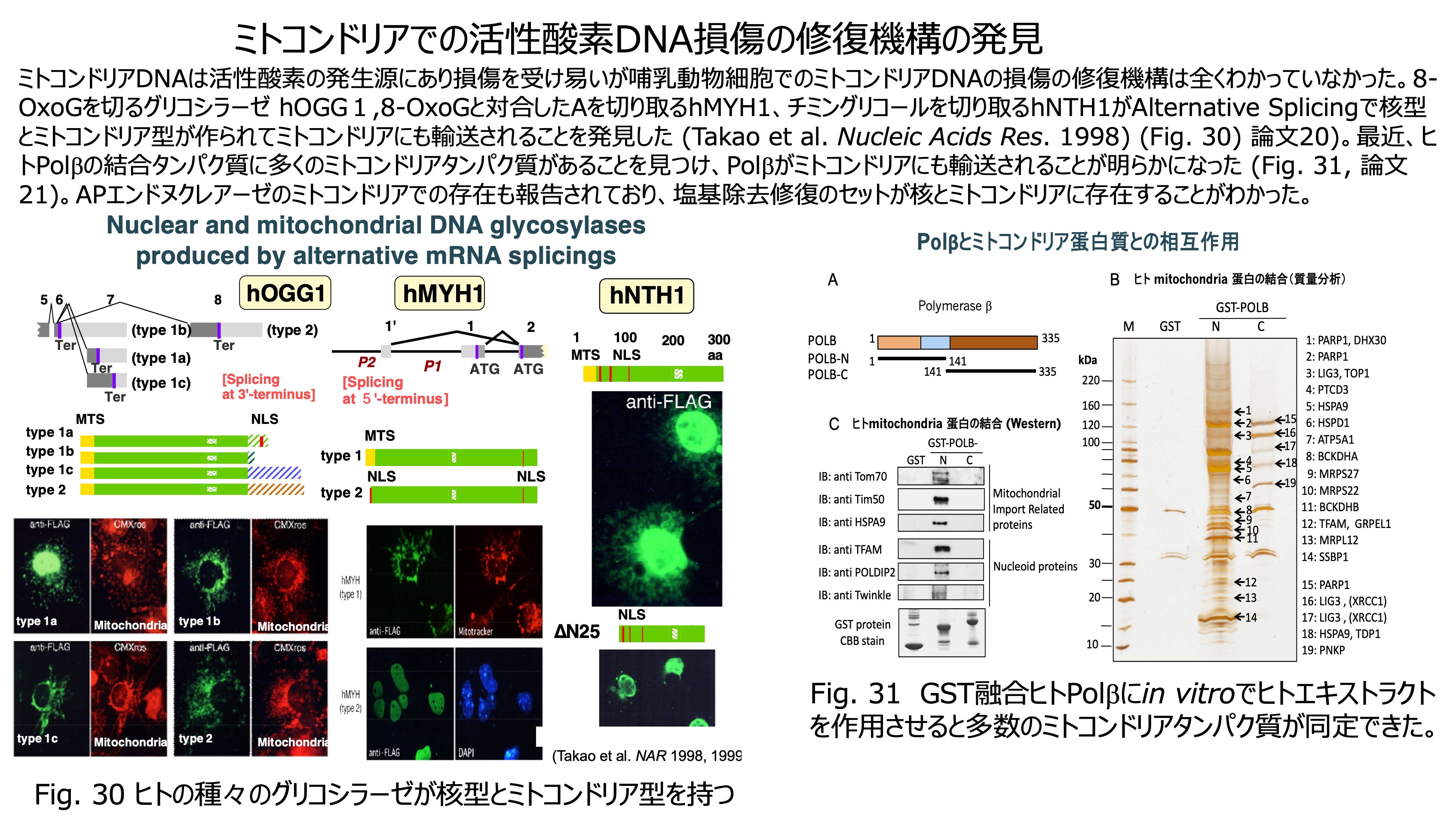
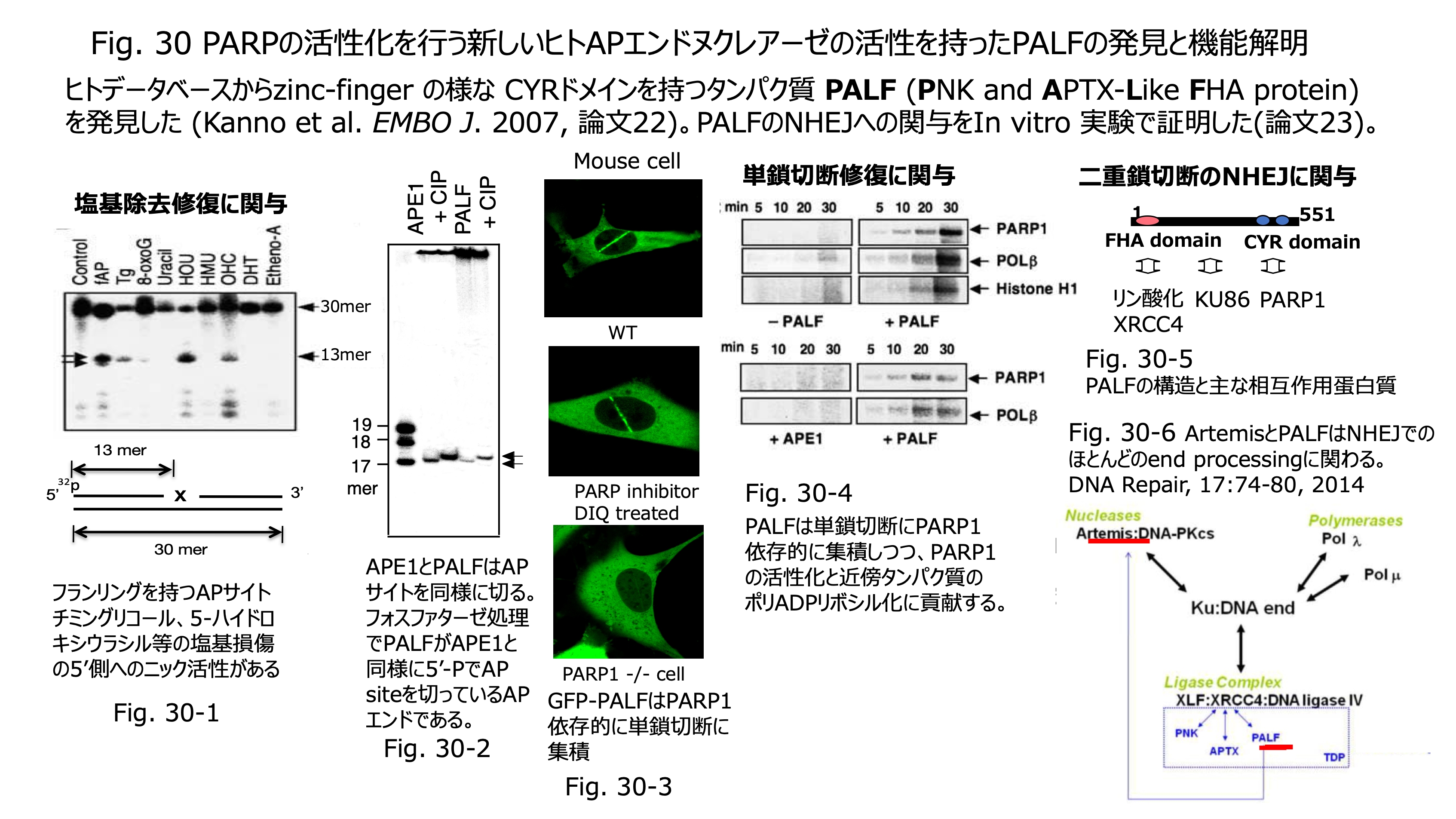
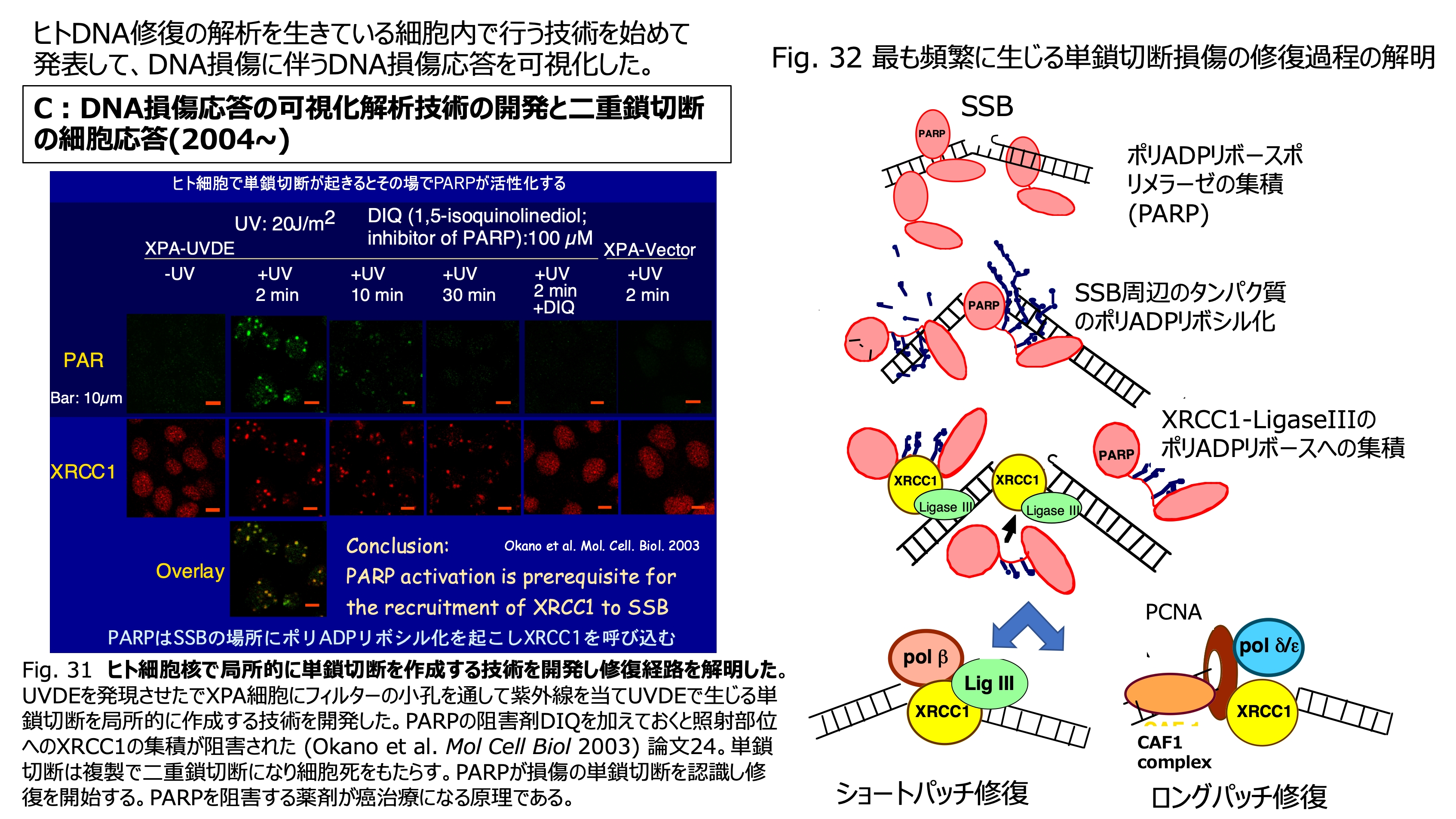
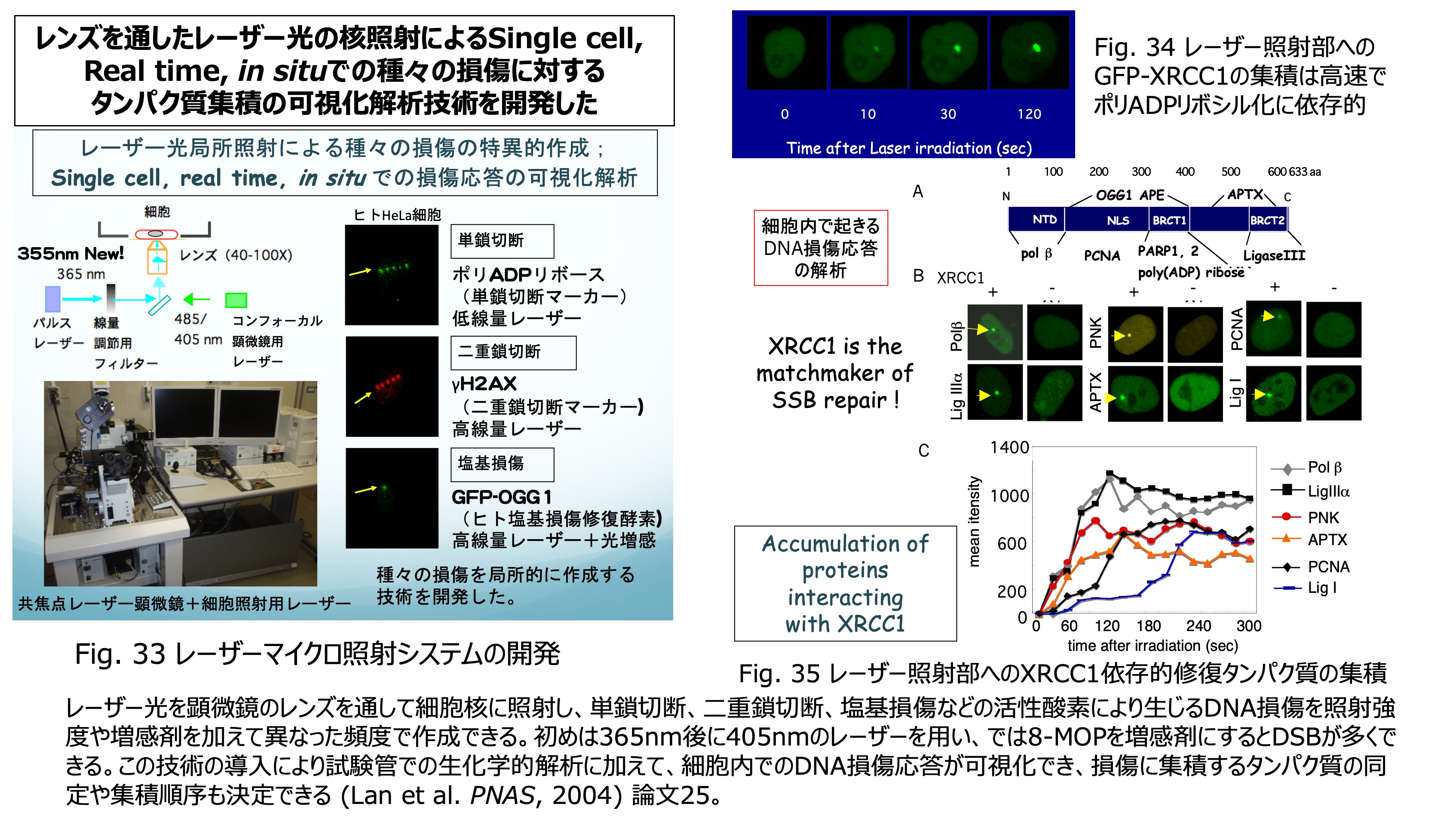
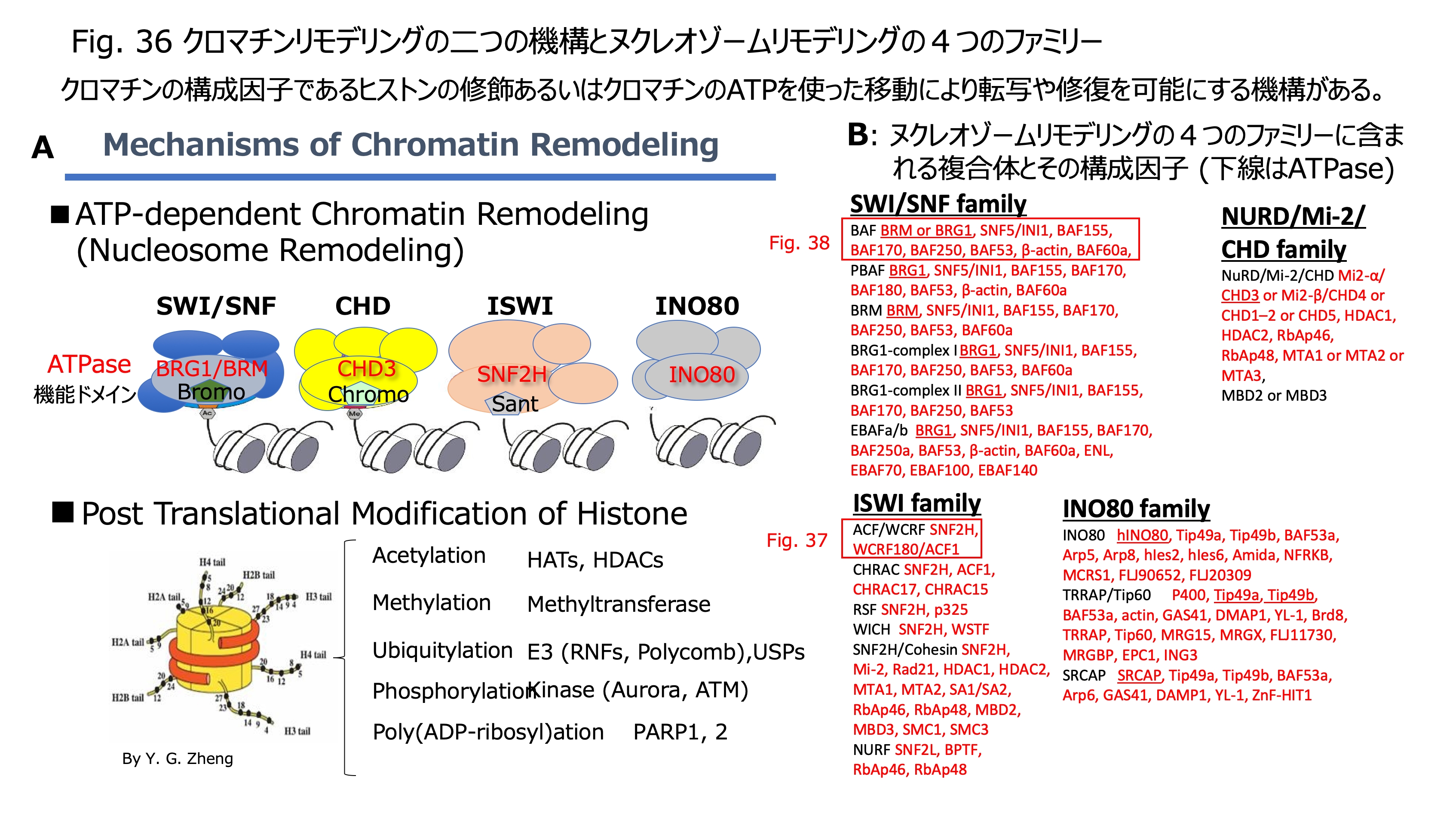
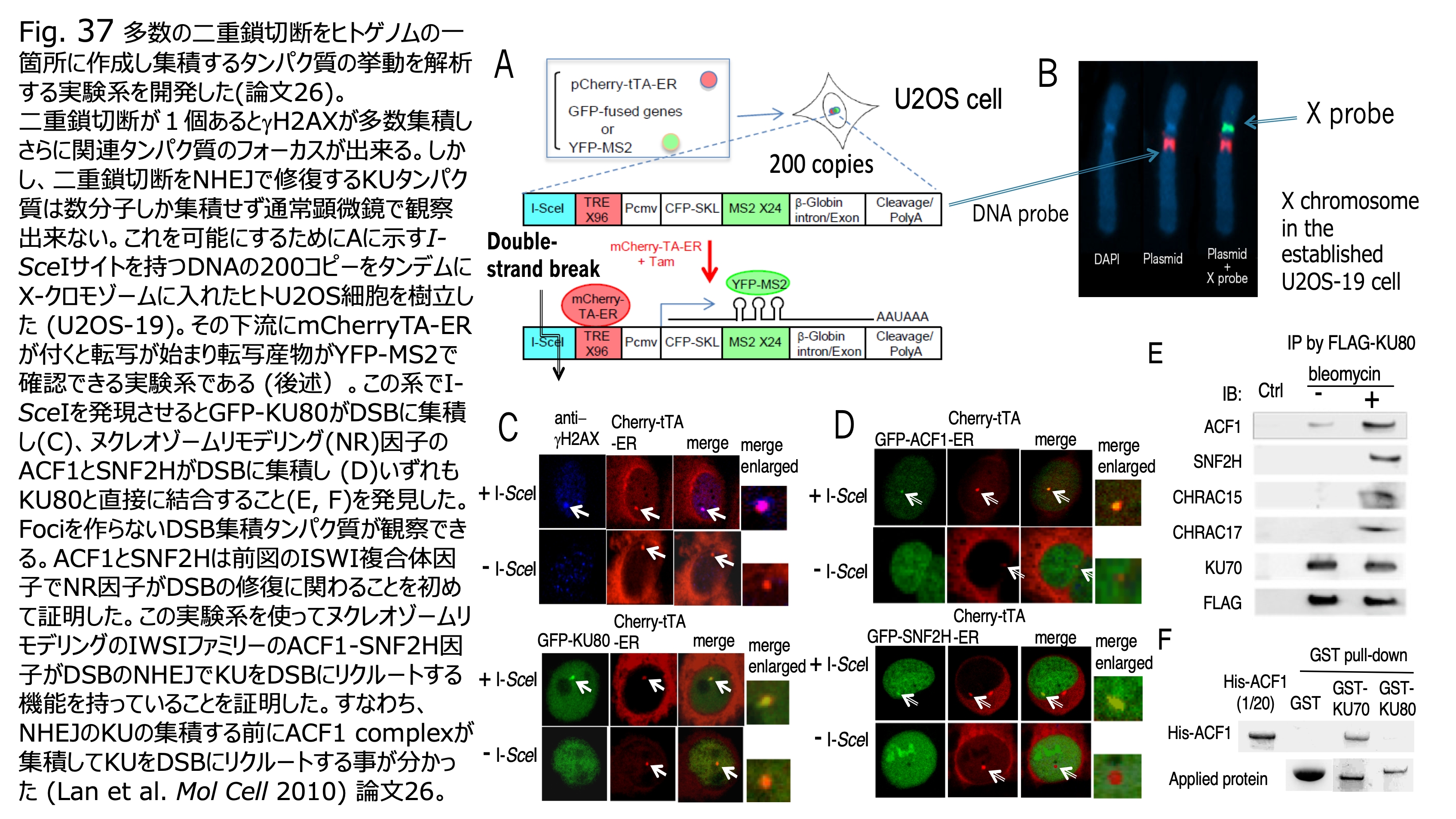
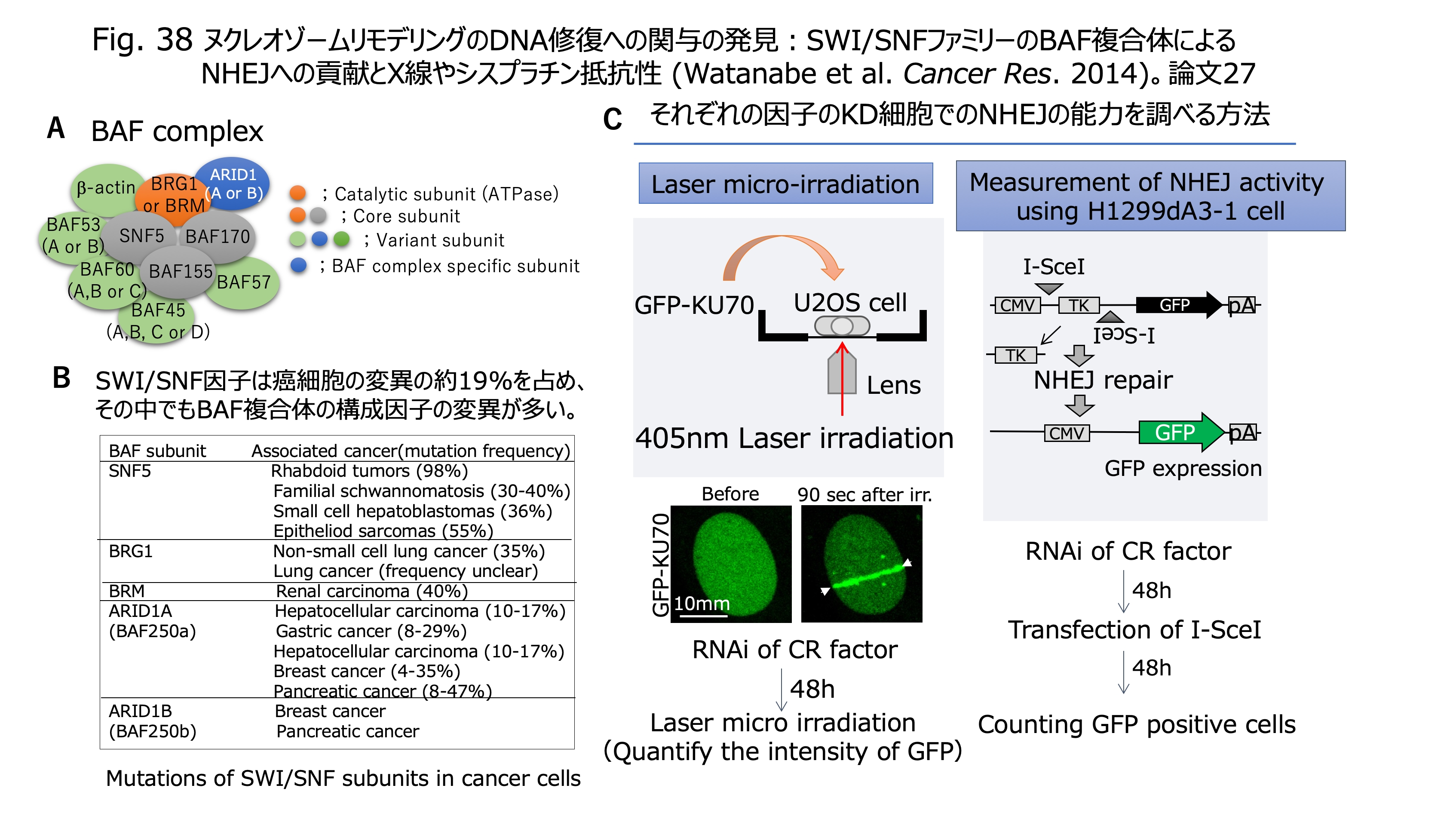
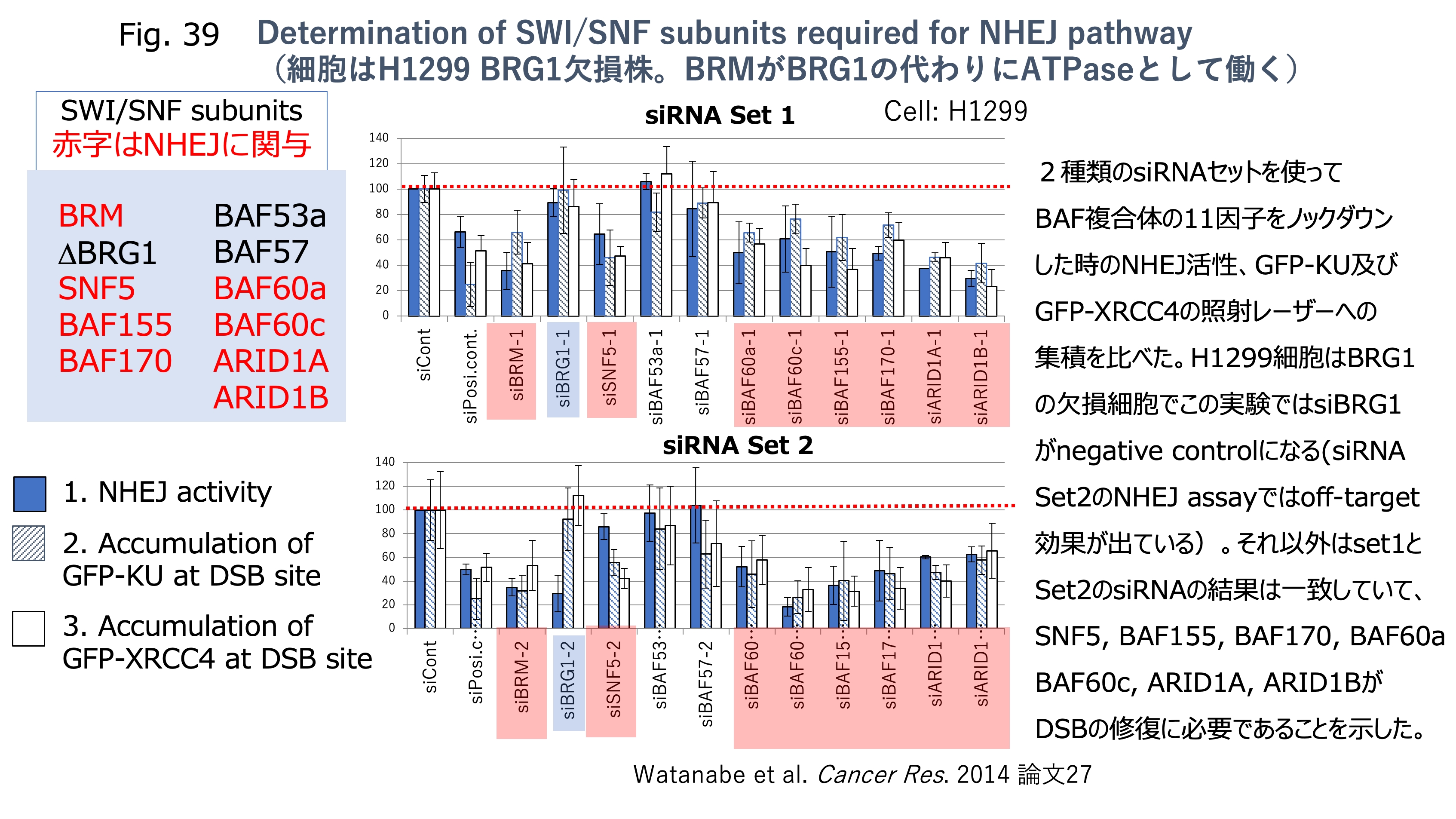
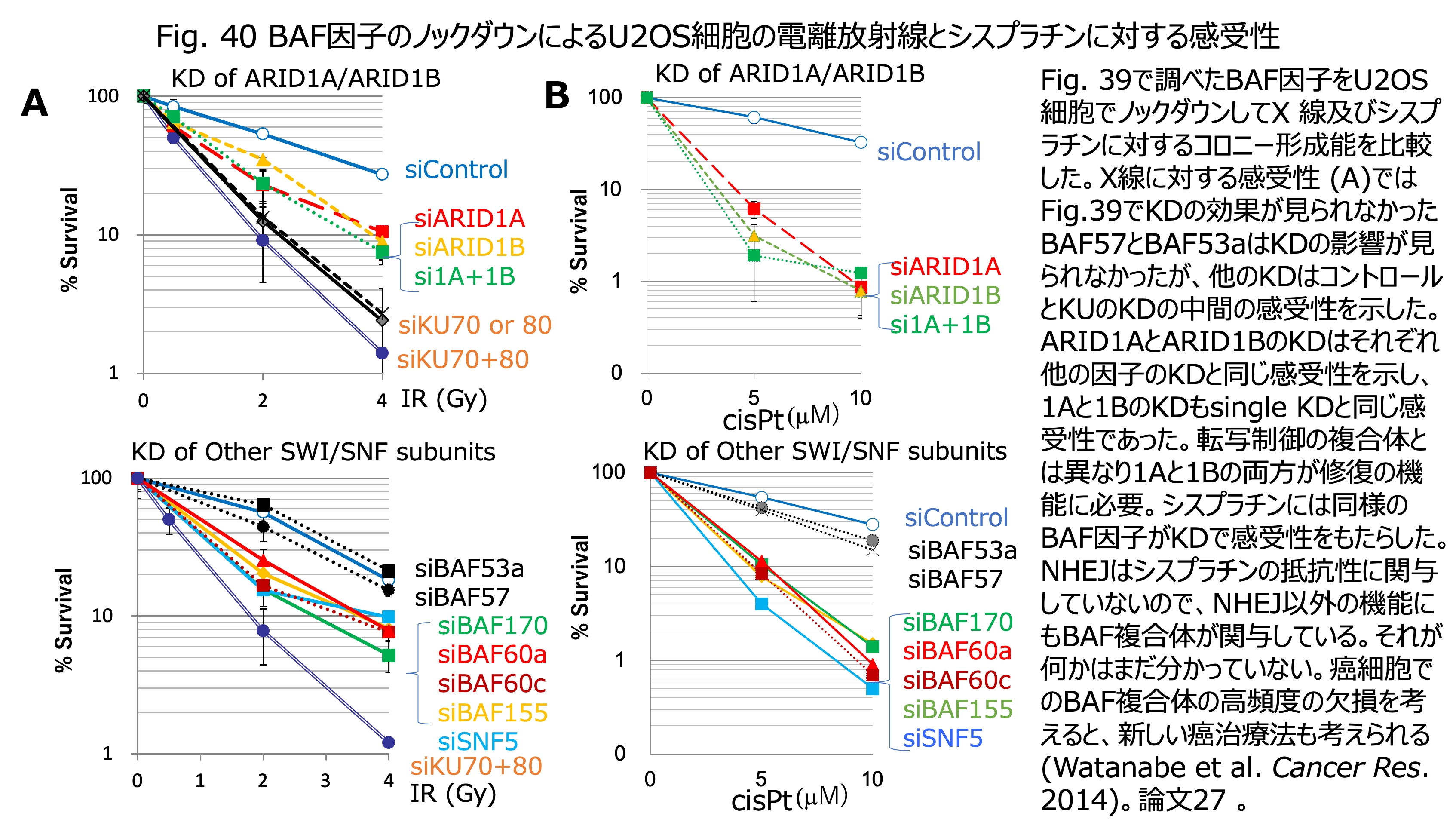
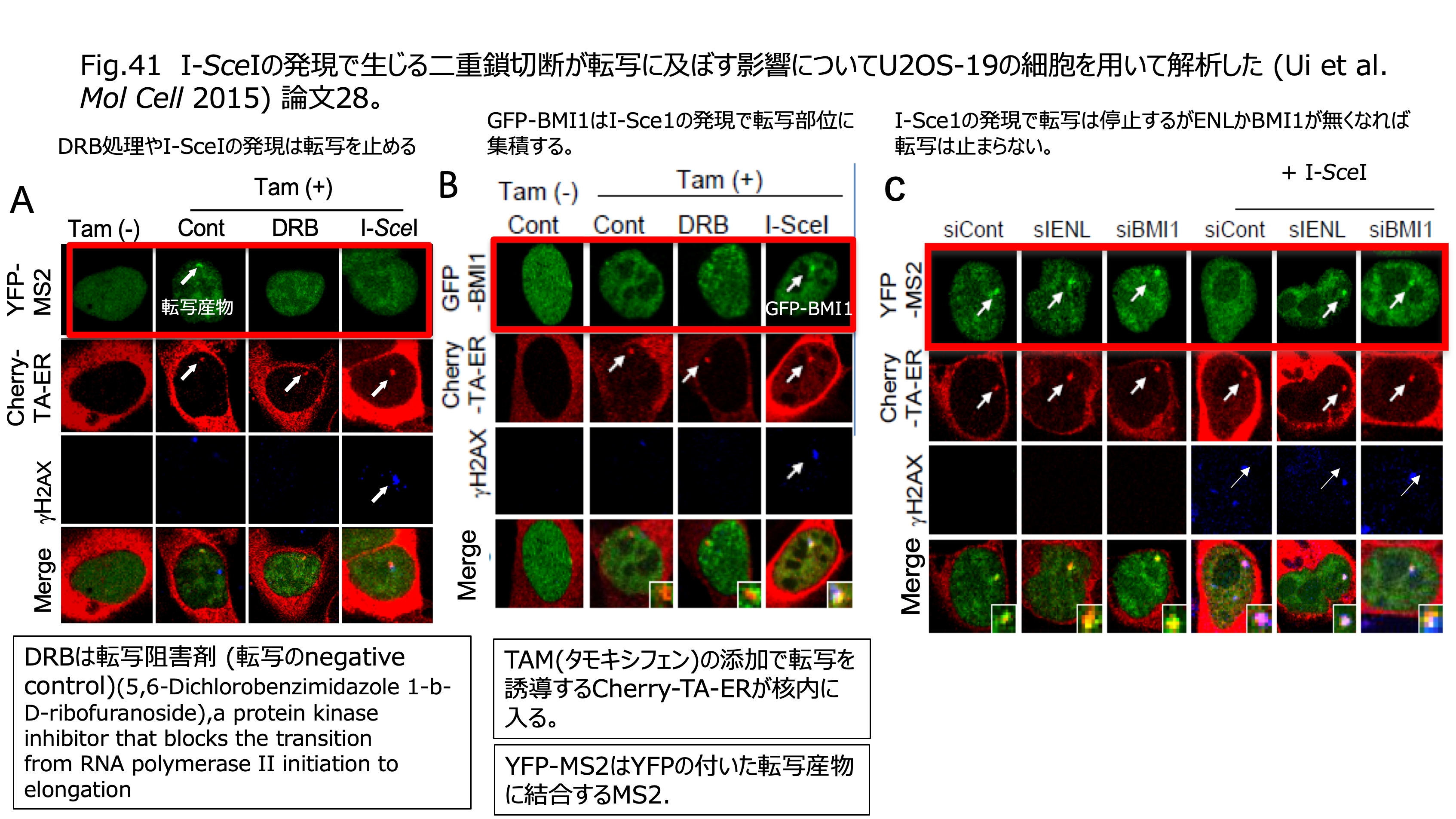
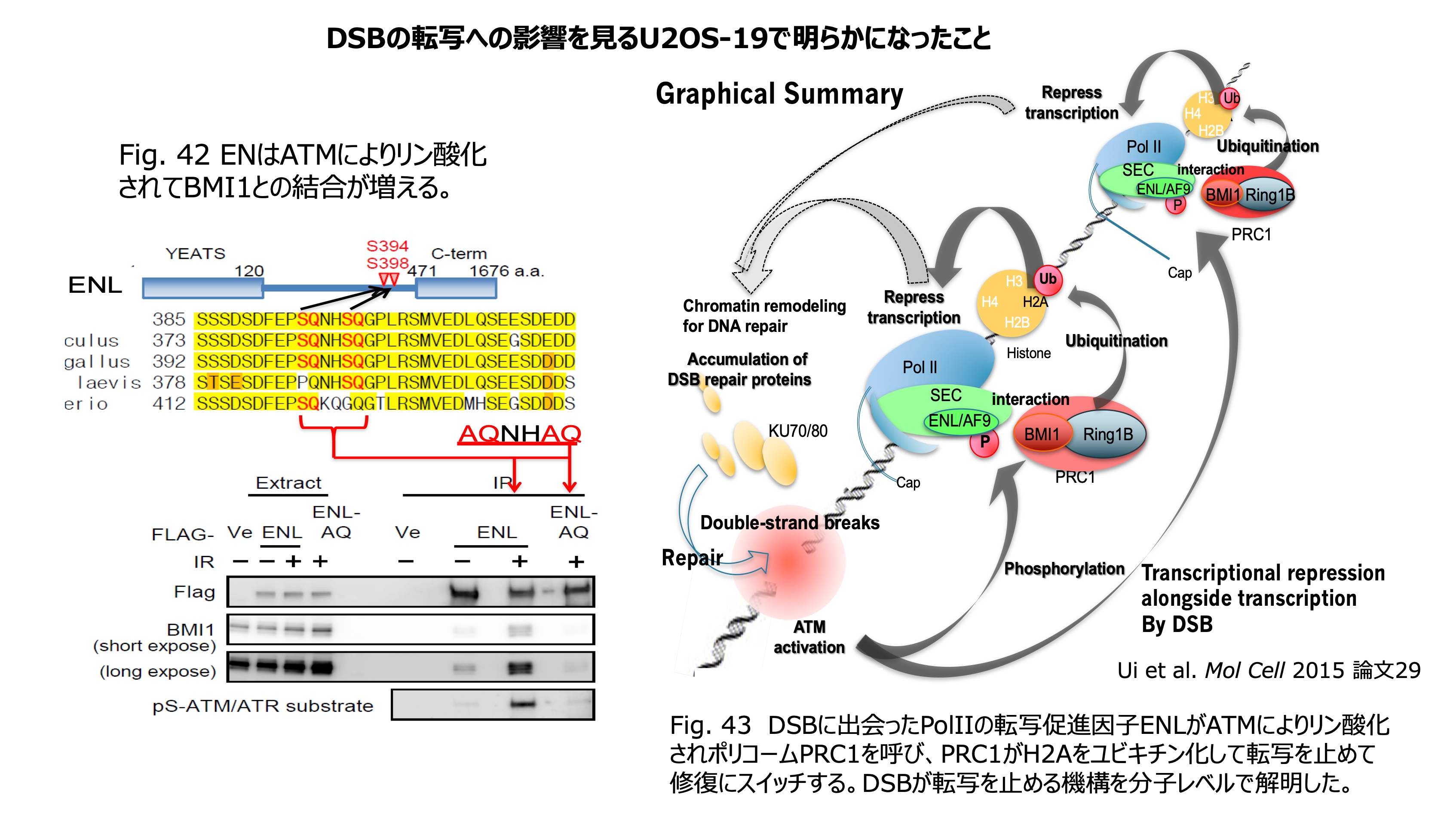
発表論文




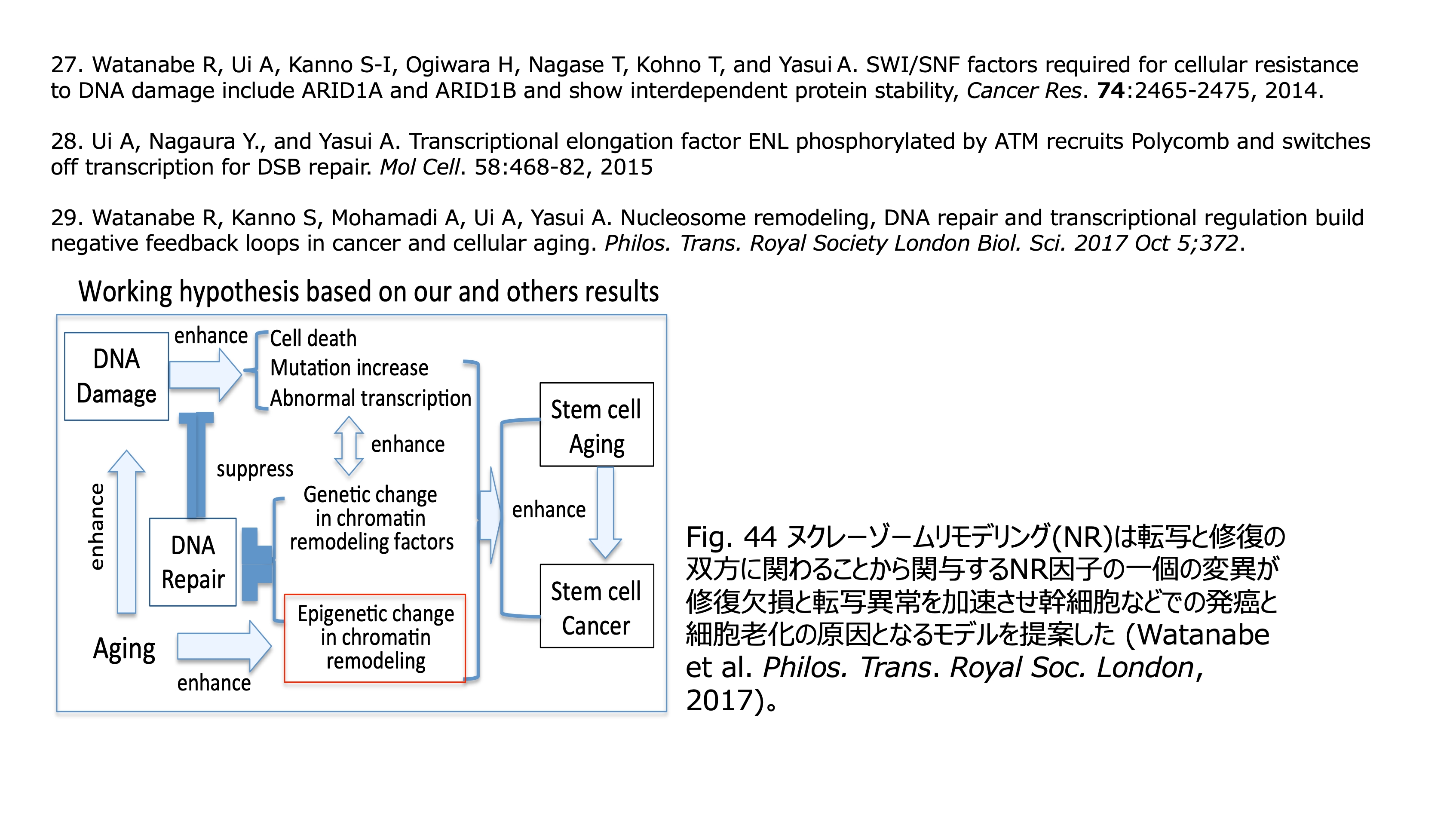

| 加齢研フェロー | 安井 明 |
平成28年4月より加齢研で新たに「フェロー」職が創設されました。この「加齢研フェロー」職は非常勤で、科研費や寄付金などの外部資金を運用しますが、科研費等の申請の際の職名として登録され、加齢研共同研究の受入教員を任され、教授会へのオブザーバー参加を許されています。事務上は特定の研究分野(分子腫瘍学)に属します。この職は加齢研の研究者や研究活動の可能性を広げることが期待されます。
研究活動
発表論文