| 教授 | 田中 耕三 |
| 助教 | 家村 顕自 |
| 助教 | 國安 絹枝 |

ゲノム不安定性が起こるしくみと、そのがんや老化との関連を解明する
私たちの体には約37兆個の細胞がありますが、これは1個の受精卵が分裂を繰り返してできたものです。この1つ1つの細胞には、精巧なしくみによって遺伝情報が正確に伝えられています。一方多くのがんでは染色体の異常や遺伝子の変異が見られ、これは遺伝情報を正確に伝えるしくみの異常(ゲノム不安定性)によって起こります。ゲノム不安定性はアルツハイマー病などの疾患や老化の過程でも見られることが報告されていますが、そのはっきりしたしくみはわかっていません。私たちはこのゲノム不安定性がどのようにして起こるのか、そしてそれがどのようにがんや老化と関連しているのかについて研究しています。そしてゲノムを安定に維持することによる疾患の予防や、ゲノム不安定性をターゲットとしたがん治療の開発につなげようとしています。
私たちは、ヒト細胞における染色体分配の分子機構について明らかにしてきました。またこの機構の異常が、どのようにして多くのがんで見られる染色体不安定性をひき起こすのかについて調べています。さらに、染色体不安定性と老化との関連についても明らかにしようとしています。一方私たちが染色体分配に関連する分子として発見したCHAMP1が、最近知的障害の原因遺伝子であることがわかったため、CHAMP1の神経系の発達への役割についても研究しています。これらの研究は、培養細胞およびマウスを用いて、ライブセルイメージング、生化学的解析、ゲノム・エピゲノム解析などの手法を駆使して行っており、分子レベルから個体レベルまでの一貫した理解を目指しています。
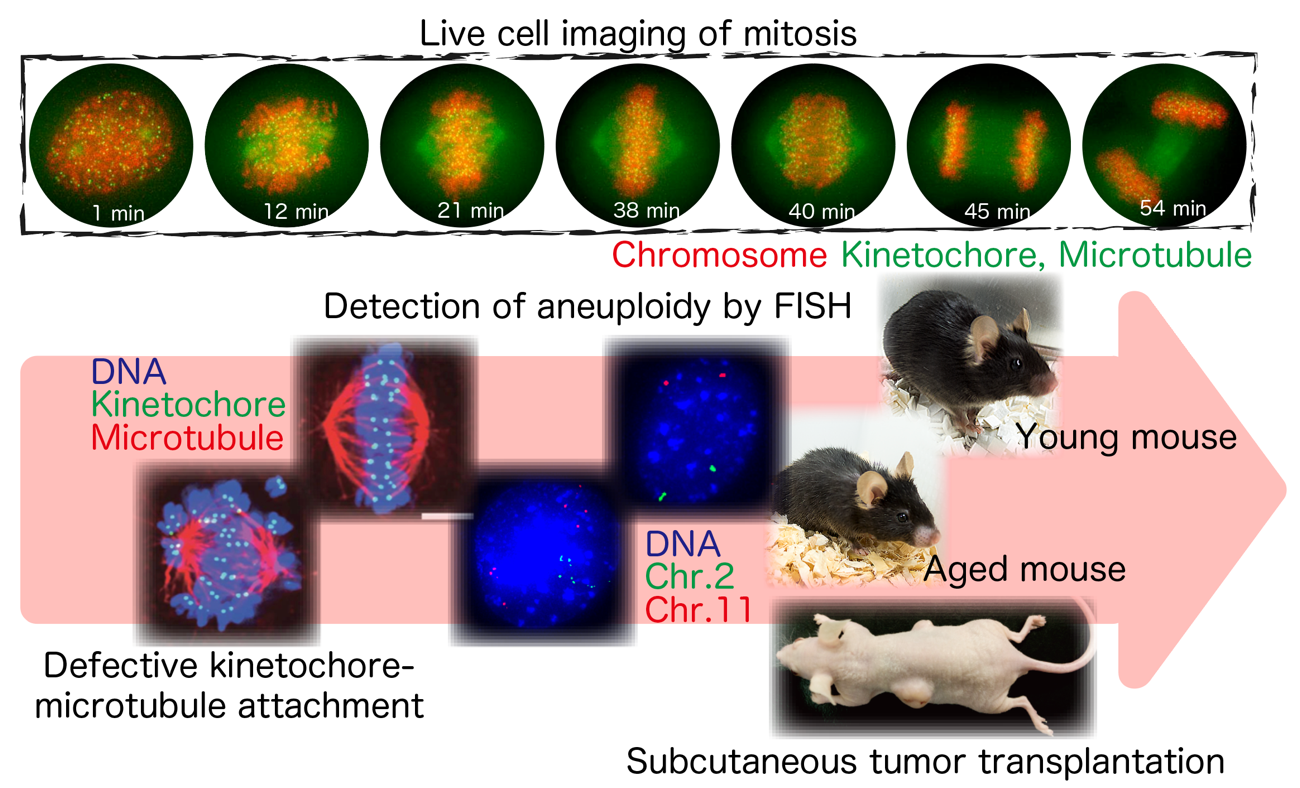
図:分子腫瘍学研究分野の研究テーマ
細胞分裂における染色体分配の過程を観察し、がんや老化で見られるゲノム不安定性がどのようにして生じるのかを研究しています。
主な研究テーマ
・がん細胞の染色体不安定性の原因の解明
・老化にともなう染色体不安定性の解明
・知的障害原因分子CHAMP1の機能の解析
・染色体不安定性とがん・老化との関連の解明
・染色体不安定性を標的としたがん治療の開発
参考文献
1. Nagai M et al. Deficiency of CHAMP1, a gene related to intellectual disability, causes impaired neuronal development and a mild behavioral phenotype. Brain Commun (2022) 4, fcac220.
2. Fujita H et al. CHAMP1-POGZ counteracts the inhibitory effect of 53BP1 on homologous recombination and affects PARP inhibitor resistance. Oncogene (2022) 41, 2706-2718.
3. Iemura K et al. Chromosome oscillation promotes Aurora A-dependent Hec1 phosphorylation and mitotic fidelity. J Cell Biol (2021) 220, e202006116.
4. Iemura, K et al. Chromokinesin Kid and kinetochore kinesin CENP-E differentially support chromosome congression without end-on attachment to microtubules. Nat Commun (2015) 6, 6447.
5. Itoh, G et al. CAMP (C13orf8, ZNF828) is a novel regulator of kinetochore-microtubule attachment. EMBO J (2011) 30, 130-144.
研究キーワード
がん、老化、染色体不安定性、ゲノム不安定性、染色体分配